In Antibody-mediated Immunity Where Does the Antigen First Bind?
The macrophages and neutrophils of the innate immune system provide a first line of defense against many common microorganisms and are essential for the control of common bacterial infections. However, they cannot always eliminate infectious organisms, and there are some pathogens that they cannot recognize. The lymphocytes of the adaptive immune system have evolved to provide a more versatile means of defense which, in addition, provides increased protection against subsequent reinfection with the same pathogen. The cells of the innate immune system, however, play a crucial part in the initiation and subsequent direction of adaptive immune responses, as well as participating in the removal of pathogens that have been targeted by an adaptive immune response. Moreover, because there is a delay of 4–7 days before the initial adaptive immune response takes effect, the innate immune response has a critical role in controlling infections during this period.
1-5. Most infectious agents induce inflammatory responses by activating innate immunity
Microorganisms such as bacteria that penetrate the epithelial surfaces of the body for the first time are met immediately by cells and molecules that can mount an innate immune response. Phagocytic macrophages conduct the defense against bacteria by means of surface receptors that are able to recognize and bind common constituents of many bacterial surfaces. Bacterial molecules binding to these receptors trigger the macrophage to engulf the bacterium and also induce the secretion of biologically active molecules. Activated macrophages secrete cytokines, which are defined as proteins released by cells that affect the behavior of other cells that bear receptors for them. They also release proteins known as chemokines that attract cells with chemokine receptors such as neutrophils and monocytes from the bloodstream (Fig. 1.12). The cytokines and chemokines released by macrophages in response to bacterial constituents initiate the process known as inflammation. Local inflammation and the phagocytosis of invading bacteria may also be triggered as a result of the activation of complement on the bacterial cell surface. Complement is a system of plasma proteins that activates a cascade of proteolytic reactions on microbial surfaces but not on host cells, coating these surfaces with fragments that are recognized and bound by phagocytic receptors on macrophages. The cascade of reactions also releases small peptides that contribute to inflammation.
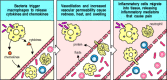
Figure 1.12
Bacterial infection triggers an inflammatory response. Macrophages encountering bacteria in the tissues are triggered to release cytokines that increase the permeability of blood vessels, allowing fluid and proteins to pass into the tissues. They also produce (more...)
Inflammation is traditionally defined by the four Latin words calor, dolor, rubor, and tumor, meaning heat, pain, redness, and swelling, all of which reflect the effects of cytokines and other inflammatory mediators on the local blood vessels. Dilation and increased permeability of the blood vessels during inflammation lead to increased local blood flow and the leakage of fluid, and account for the heat, redness, and swelling. Cytokines and complement fragments also have important effects on the adhesive properties of the endothelium, causing circulating leukocytes to stick to the endothelial cells of the blood vessel wall and migrate between them to the site of infection, to which they are attracted by chemokines. The migration of cells into the tissue and their local actions account for the pain. The main cell types seen in an inflammatory response in its initial phases are neutrophils, which are recruited into the inflamed, infected tissue in large numbers. Like macrophages, they have surface receptors for common bacterial constituents and complement, and they are the principal cells that engulf and destroy the invading micro-organisms. The influx of neutrophils is followed a short time later by monocytes that rapidly differentiate into macrophages. Macrophages and neutrophils are thus also known as inflammatory cells. Inflammatory responses later in an infection also involve lymphocytes, which have meanwhile been activated by antigen that has drained from the site of infection via the afferent lymphatics.
The innate immune response makes a crucial contribution to the activation of adaptive immunity. The inflammatory response increases the flow of lymph containing antigen and antigen-bearing cells into lymphoid tissue, while complement fragments on microbial surfaces and induced changes in cells that have taken up microorganisms provide signals that synergize in activating lymphocytes whose receptors bind to specific microbial antigens. Macrophages that have phagocytosed bacteria and become activated can also activate T lymphocytes. However, the cells that specialize in presenting antigen to T lymphocytes and initiating adaptive immunity are the dendritic cells.
1-6. Activation of specialized antigen-presenting cells is a necessary first step for induction of adaptive immunity
The induction of an adaptive immune response begins when a pathogen is ingested by an immature dendritic cell in the infected tissue. These specialized phagocytic cells are resident in most tissues and are relatively long-lived, turning over at a slow rate. They derive from the same bone marrow precursor as macrophages, and migrate from the bone marrow to their peripheral stations, where their role is to survey the local environment for pathogens. Eventually, all tissue-resident dendritic cells migrate through the lymph to the regional lymph nodes where they interact with recirculating naive lymphocytes. If the dendritic cells fail to be activated, they induce tolerance to the antigens of self that they bear.
The immature dendritic cell carries receptors on its surface that recognize common features of many pathogens, such as bacterial cell wall proteoglycans. As with macrophages and neutrophils, binding of a bacterium to these receptors stimulates the dendritic cell to engulf the pathogen and degrade it intracellularly. Immature dendritic cells are also continually taking up extracellular material, including any virus particles or bacteria that may be present, by the receptor-independent mechanism of macropinocytosis. The function of dendritic cells, however, is not primarily to destroy pathogens but to carry pathogen antigens to peripheral lymphoid organs and there present them to T lymphocytes. When a dendritic cell takes up a pathogen in infected tissue, it becomes activated, and travels to a nearby lymph node. On activation, the dendritic cell matures into a highly effective antigen-presenting cell (APC) and undergoes changes that enable it to activate pathogen-specific lymphocytes that it encounters in the lymph node (Fig. 1.13). Activated dendritic cells secrete cytokines that influence both innate and adaptive immune responses, making these cells essential gatekeepers that determine whether and how the immune system responds to the presence of infectious agents. We shall consider the maturation of dendritic cells and their central role in presenting antigens to T lymphocytes in Chapter 8.

Figure 1.13
Dendritic cells initiate adaptive immune responses. Immature dendritic cells resident in infected tissues take up pathogens and their antigens by macropinocytosis and receptor-mediated phagocytosis. They are stimulated by recognition of the presence of (more...)
1-7. Lymphocytes activated by antigen give rise to clones of antigen-specific cells that mediate adaptive immunity
The defense systems of innate immunity are effective in combating many pathogens. They are constrained, however, by relying on germline-encoded receptors to recognize microorganisms that can evolve more rapidly than the hosts they infect. This explains why they can only recognize microorganisms bearing surface molecules that are common to many pathogens and that have been conserved over the course of evolution. Not surprisingly, many pathogenic bacteria have evolved a protective capsule that enables them to conceal these molecules and thereby avoid being recognized and phagocytosed. Viruses carry no invariant molecules similar to those of bacteria and are rarely recognized directly by macrophages. Viruses and encapsulated bacteria can, however, still be taken up by dendritic cells through the nonreceptor-dependent process of macropinocytosis. Molecules that reveal their infectious nature may then be unmasked, and the dendritic cell activated to present their antigens to lymphocytes. The recognition mechanism used by the lymphocytes of the adaptive immune response has evolved to overcome the constraints faced by the innate immune system, and enables recognition of an almost infinite diversity of antigens, so that each different pathogen can be targeted specifically.
Instead of bearing several different receptors, each recognizing a different surface feature shared by many pathogens, each naive lymphocyte entering the bloodstream bears antigen receptors of a single specificity. The specificity of these receptors is determined by a unique genetic mechanism that operates during lymphocyte development in the bone marrow and thymus to generate millions of different variants of the genes encoding the receptor molecules. Thus, although an individual lymphocyte carries receptors of only one specificity, the specificity of each lymphocyte is different. This ensures that the millions of lymphocytes in the body collectively carry millions of different antigen receptor specificities—the lymphocyte receptor repertoire of the individual. During a person's lifetime these lymphocytes undergo a process akin to natural selection; only those lymphocytes that encounter an antigen to which their receptor binds will be activated to proliferate and differentiate into effector cells.
This selective mechanism was first proposed in the 1950s by Macfarlane Burnet to explain why antibodies, which can be induced in response to virtually any antigen, are produced in each individual only to those antigens to which he or she is exposed. He postulated the preexistence in the body of many different potential antibody-producing cells, each having the ability to make antibody of a different specificity and displaying on its surface a membrane-bound version of the antibody that served as a receptor for antigen. On binding antigen, the cell is activated to divide and produce many identical progeny, known as a clone; these cells can now secrete clonotypic antibodies with a specificity identical to that of the surface receptor that first triggered activation and clonal expansion (Fig. 1.14). Burnet called this the clonal selection theory.
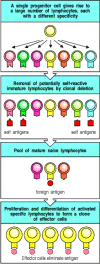
Figure 1.14
Clonal selection. Each lymphocyte progenitor gives rise to many lymphocytes, each bearing a distinct antigen receptor. Lymphocytes with receptors that bind ubiquitous self antigens are eliminated before they become fully mature, ensuring tolerance to (more...)
1-8. Clonal selection of lymphocytes is the central principle of adaptive immunity
Remarkably, at the time that Burnet formulated his theory, nothing was known of the antigen receptors of lymphocytes; indeed the function of lymphocytes themselves was still obscure. Lymphocytes did not take center stage until the early 1960s, when James Gowans discovered that removal of the small lymphocytes from rats resulted in the loss of all known adaptive immune responses. These immune responses were restored when the small lymphocytes were replaced. This led to the realization that lymphocytes must be the units of clonal selection, and their biology became the focus of the new field of cellular immunology.
Clonal selection of lymphocytes with diverse receptors elegantly explained adaptive immunity but it raised one significant intellectual problem. If the antigen receptors of lymphocytes are generated randomly during the lifetime of an individual, how are lymphocytes prevented from recognizing antigens on the tissues of the body and attacking them? Ray Owen had shown in the late 1940s that genetically different twin calves with a common placenta were immunologically tolerant of one another's tissues, that is, they did not make an immune response against each other. Sir Peter Medawar then showed in 1953 that if exposed to foreign tissues during embryonic development, mice become immunologically tolerant to these tissues. Burnet proposed that developing lymphocytes that are potentially self-reactive are removed before they can mature, a process known as clonal deletion. He has since been proved right in this too, although the mechanisms of tolerance are still being worked out, as we shall see when we discuss the development of lymphocytes in Chapter 7.
Clonal selection of lymphocytes is the single most important principle in adaptive immunity. Its four basic postulates are listed in Fig. 1.15. The last of the problems posed by the clonal selection theory—how the diversity of lymphocyte antigen receptors is generated—was solved in the 1970s when advances in molecular biology made it possible to clone the genes encoding antibody molecules.

Figure 1.15
The four basic principles of clonal selection.
1-9. The structure of the antibody molecule illustrates the central puzzle of adaptive immunity
Antibodies, as discussed above, are the secreted form of the B-cell antigen receptor or BCR. Because they are produced in very large quantities in response to antigen, they can be studied by traditional biochemical techniques; indeed, their structure was understood long before recombinant DNA technology made it possible to study the membrane-bound antigen receptors of lymphocytes. The startling feature that emerged from the biochemical studies was that an antibody molecule is composed of two distinct regions. One is a constant region that can take one of only four or five biochemically distinguishable forms; the other is a variable region that can take an apparently infinite variety of subtly different forms that allow it to bind specifically to an equally vast variety of different antigens.
This division is illustrated in the simple schematic diagram in Fig. 1.16, where the antibody is depicted as a Y-shaped molecule, with the constant region shown in blue and the variable region in red. The two variable regions, which are identical in any one antibody molecule, determine the antigen-binding specificity of the antibody; the constant region determines how the antibody disposes of the pathogen once it is bound.
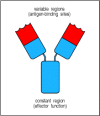
Figure 1.16
Schematic structure of an antibody molecule. The two arms of the Y-shaped antibody molecule contain the variable regions that form the two identical antigen-binding sites. The stem can take one of only a limited number of forms and is known as the constant (more...)
Each antibody molecule has a twofold axis of symmetry and is composed of two identical heavy chains and two identical light chains (Fig. 1.17). Heavy and light chains both have variable and constant regions; the variable regions of a heavy and a light chain combine to form an antigen-binding site, so that both chains contribute to the antigen-binding specificity of the antibody molecule. The structure of antibody molecules will be described in detail in Chapter 3, and the functional properties of antibodies conferred by their constant regions will be considered in Chapters 4 and 9. For the time being we are concerned only with the properties of immunoglobulin molecules as antigen receptors, and how the diversity of the variable regions is generated.

Figure 1.17
Antibodies are made up of four protein chains. There are two types of chain in an antibody molecule: a larger chain called the heavy chain (green), and a smaller one called the light chain (yellow). Each chain has both a variable and a constant region, (more...)
1-10. Each developing lymphocyte generates a unique antigen receptor by rearranging its receptor genes
How are antigen receptors with an almost infinite range of specificities encoded by a finite number of genes? This question was answered in 1976, when Susumu Tonegawa discovered that the genes for immunoglobulin variable regions are inherited as sets of gene segments, each encoding a part of the variable region of one of the immunoglobulin polypeptide chains (Fig. 1.18). During B-cell development in the bone marrow, these gene segments are irreversibly joined by DNA recombination to form a stretch of DNA encoding a complete variable region. Because there are many different gene segments in each set, and different gene segments are joined together in different cells, each cell generates unique genes for the variable regions of the heavy and light chains of the immunoglobulin molecule. Once these recombination events have succeeded in producing a functional receptor, further rearrangement is prohibited. Thus each lymphocyte expresses only one receptor specificity.
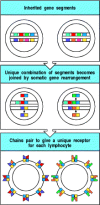
Figure 1.18
The diversity of lymphocyte antigen receptors is generated by somatic gene rearrangements. Different parts of the variable regions of antigen receptors are encoded by sets of gene segments. During a lymphocyte's development, one member of each set of (more...)
This mechanism has three important consequences. First, it enables a limited number of gene segments to generate a vast number of different proteins. Second, because each cell assembles a different set of gene segments, each cell expresses a unique receptor specificity. Third, because gene rearrangement involves an irreversible change in a cell's DNA, all the progeny of that cell will inherit genes encoding the same receptor specificity. This general scheme was later also confirmed for the genes encoding the antigen receptor on T lymphocytes. The main distinctions between B- and T-lymphocyte receptors are that the immunoglobulin that serves as the B-cell antigen receptor has two identical antigen-recognition sites and can also be secreted, whereas the T-cell antigen receptor has a single antigen-recognition site and is always a cell-surface molecule. We shall see later that these receptors also recognize antigen in very different ways.
The potential diversity of lymphocyte receptors generated in this way is enormous. Just a few hundred different gene segments can combine in different ways to generate thousands of different receptor chains. The diversity of lymphocyte receptors is further amplified by junctional diversity, created by adding or subtracting nucleotides in the process of joining the gene segments, and by the fact that each receptor is made by pairing two different variable chains, each encoded in distinct sets of gene segments. A thousand different chains of each type could thus generate 106 distinct antigen receptors through this combinatorial diversity. Thus a small amount of genetic material can encode a truly staggering diversity of receptors. Only a subset of these randomly generated receptor specificities survive the selective processes that shape the peripheral lymphocyte repertoire; nevertheless, there are lymphocytes of at least 108 different specificities in an individual at any one time. These provide the raw material on which clonal selection acts.
1-11. Lymphocyte development and survival are determined by signals received through their antigen receptors
Equally amazing as the generation of millions of specificities of lymphocyte antigen receptors is the shaping of this repertoire during lymphocyte development and the homeostatic maintenance of such an extensive repertoire in the periphery. How are the most useful receptor specificities selected, and how are the numbers of peripheral lymphocytes, and the percentages of B cells and T cells kept relatively constant? The answer seems to be that lymphocyte maturation and survival are regulated by signals received through their antigen receptors. Strong signals received through the antigen receptor by an immature lymphocyte cause it to die or undergo further receptor rearrangement, and in this way self-reactive receptor specificities are deleted from the repertoire. However, a complete absence of signals from the antigen receptor can also lead to cell death. It seems that in order to survive, lymphocytes must periodically receive certain signals from their environment via their antigen receptors. In this way, the body can ensure that each receptor is functional and regulate the number and type of lymphocytes in the population at any given time. These survival signals appear to be delivered by other cells in the lymphoid organs and must derive, at least in part, from the body's own molecules, the self antigens, as altering the self environment alters the life-span of lymphocytes in that environment. Developing B cells in the bone marrow interact with stromal cells, while their final maturation and continued recirculation appears to depend on survival signals received from the B-cell follicles of peripheral lymphoid tissue. T lymphocytes receive survival signals from self molecules on specialized epithelial cells in the thymus during development, and from the same molecules expressed by dendritic cells in the lymphoid tissues in the periphery. The self ligands that interact with the T-cell receptor to deliver these signals are partially defined, being composed of known cell-surface molecules complexed with undefined peptides from other self proteins in the cell. These same cell-surface molecules function to present foreign intracellular antigens to T cells, as we shall explain in Section 1-16, and in Chapter 5. They select only a subset of T-cell receptors for survival, but these are the receptors most likely to be useful in responding to foreign antigens, as we shall see in Chapter 7.
Lymphocytes that fail to receive survival signals, and those that are clonally deleted because they are self-reactive, undergo a form of cell suicide called apoptosis or programmed cell death. Apoptosis, derived from a Greek word meaning the falling of leaves from the trees, occurs in all tissues, at a relatively constant rate in each tissue, and is a means of regulating the number of cells in the body. It is responsible, for example, for the death and shedding of skin cells, the turnover of liver cells, and the death of the oldest intestinal epithelial cells that are constantly replaced by new cells. Thus, it should come as no surprise that immune system cells are regulated through the same mechanism. Each day the bone marrow produces many millions of new neutrophils, monocytes, red blood cells, and lymphocytes, and this production must be balanced by an equal loss of these cells. Regulated loss of all these blood cells occurs by apoptosis, and the dying cells are finally phagocytosed by specialized macrophages in the liver and spleen. Lymphocytes are a special case, because the loss of an individual naive lymphocyte means the loss of a receptor specificity from the repertoire, while each newly matured cell that survives will contribute a different specificity. The survival signals received through the antigen receptors appear to regulate this process by inhibiting the apoptosis of individual lymphocytes, thus regulating the maintenance and composition of the lymphocyte repertoire. We shall return to the question of which ligands deliver these signals, and how they contribute to shaping and maintaining the receptor repertoire, in Chapter 7.
1-12. Lymphocytes proliferate in response to antigen in peripheral lymphoid organs, generating effector cells and immunological memory
The large diversity of lymphocyte receptors means that there will usually be at least a few that can bind to any given foreign antigen. However, because each lymphocyte has a different receptor, the numbers of lymphocytes that can bind and respond to any given antigen is very small. To generate sufficient antigen-specific effector lymphocytes to fight an infection, a lymphocyte with an appropriate receptor specificity must be activated to proliferate before its progeny finally differentiate into effector cells. This clonal expansion is a feature common to all adaptive immune responses.
As we have seen, lymphocyte activation and proliferation is initiated in the draining lymphoid tissues, where naive lymphocytes and activated antigen-presenting cells can come together. Antigens are thus presented to the naive recirculating lymphocytes as they migrate through the lymphoid tissue before returning to the bloodstream via the efferent lymph. On recognizing its specific antigen, a small lymphocyte stops migrating and enlarges. The chromatin in its nucleus becomes less dense, nucleoli appear, the volume of both the nucleus and the cytoplasm increases, and new RNAs and proteins are synthesized. Within a few hours, the cell looks completely different and is known as a lymphoblast (Fig. 1.19).

Figure 1.19
Transmission electron micrographs of lymphocytes at various stages of activation to effector function. Small resting lymphocytes (top panel) have not yet encountered antigen. Note the scanty cytoplasm, the absence of rough endoplasmic reticulum, and the (more...)
The lymphoblasts now begin to divide, normally duplicating themselves two to four times every 24 hours for 3 to 5 days, so that one naive lymphocyte gives rise to a clone of around 1000 daughter cells of identical specificity. These then differentiate into effector cells (see Fig. 1.19). In the case of B cells, the differentiated effector cells, the plasma cells, secrete antibody; in the case of T cells, the effector cells are able to destroy infected cells or activate other cells of the immune system. These changes also affect the recirculation of antigen-specific lymphocytes. Changes in the cell-adhesion molecules they express on their surface allow effector lymphocytes to migrate into sites of infection or stay in the lymphoid organs to activate B cells.
After a naive lymphocyte has been activated, it takes 4 to 5 days before clonal expansion is complete and the lymphocytes have differentiated into effector cells. That is why adaptive immune responses occur only after a delay of several days. Effector cells have only a limited life-span and, once antigen is removed, most of the antigen-specific cells generated by the clonal expansion of small lymphocytes undergo apoptosis. However, some persist after the antigen has been eliminated. These cells are known as memory cells and form the basis of immunological memory, which ensures a more rapid and effective response on a second encounter with a pathogen and thereby provides lasting protective immunity.
The characteristics of immunological memory are readily observed by comparing the antibody response of an individual to a first or primary immunization with the response elicited in the same individual by a secondary or booster immunization with the same antigen. As shown in Fig. 1.20, the secondary antibody response occurs after a shorter lag phase, achieves a markedly higher level, and produces antibodies of higher affinity, or strength of binding, for the antigen. We shall describe the mechanisms of these remarkable changes in Chapters 9 and 10. The cellular basis of immunological memory is the clonal expansion and clonal differentiation of cells specific for the eliciting antigen, and it is therefore entirely antigen specific.
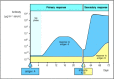
Figure 1.20
The course of a typical antibody response. First encounter with an antigen produces a primary response. Antigen A introduced at time zero encounters little specific antibody in the serum. After a lag phase, antibody against antigen A (blue) appears; its (more...)
It is immunological memory that enables successful vaccination and prevents reinfection with pathogens that have been repelled successfully by an adaptive immune response. Immunological memory is the most important biological consequence of the development of adaptive immunity, although its cellular and molecular basis is still not fully understood, as we shall see in Chapter 10.
1-13. Interaction with other cells as well as with antigen is necessary for lymphocyte activation
Peripheral lymphoid tissues are specialized not only to trap phagocytic cells that have ingested antigen (see Sections 1-3 and 1-6) but also to promote their interactions with lymphocytes that are needed to initiate an adaptive immune response. The spleen and lymph nodes in particular are highly organized for the latter function.
All lymphocyte responses to antigen require not only the signal that results from antigen binding to their receptors, but also a second signal, which is delivered by another cell. Naive T cells are generally activated by activated dendritic cells (Fig. 1.21, left panel) but for B cells (Fig. 1.21, right panel), the second signal is delivered by an armed effector T cell. Because of their ability to deliver activating signals, these three cell types are known as professional antigen-presenting cells, or often just antigen-presenting cells. They are illustrated in Fig. 1.22. Dendritic cells are the most important antigenpresenting cell of the three, with a central role in the initiation of adaptive immune responses (see Section 1-6). Macrophages can also mediate innate immune responses directly and make a crucial contribution to the effector phase of the adaptive immune response. B cells contribute to adaptive immunity by presenting peptides from antigens they have ingested and by secreting antibody.

Figure 1.21
Two signals are required for lymphocyte activation. As well as receiving a signal through their antigen receptor, mature naive lymphocytes must also receive a second signal to become activated. For T cells (left panel) it is delivered by a professional (more...)
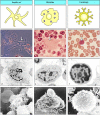
Figure 1.22
The professional antigen-presenting cells. The three types of professional antigen-presenting cell are shown in the form in which they will be depicted throughout this book (top row), as they appear in the light microscope (second row; the relevant cell (more...)
Thus, the final postulate of adaptive immunity is that it occurs on a cell that also presents the antigen. This appears to be an absolute rule in vivo, although exceptions have been observed in in vitro systems. Nevertheless, what we are attempting to define is what does happen, not what can happen.
Summary
The early innate systems of defense, which depend on invariant receptors recognizing common features of pathogens, are crucially important, but they are evaded or overcome by many pathogens and do not lead to immunological memory. The abilities to recognize all pathogens specifically and to provide enhanced protection against reinfection are the unique features of adaptive immunity, which is based on clonal selection of lymphocytes bearing antigen-specific receptors. The clonal selection of lymphocytes provides a theoretical framework for understanding all the key features of adaptive immunity. Each lymphocyte carries cell-surface receptors of a single specificity, generated by the random recombination of variable receptor gene segments and the pairing of different variable chains. This produces lymphocytes, each bearing a distinct receptor, so that the total repertoire of receptors can recognize virtually any antigen. If the receptor on a lymphocyte is specific for a ubiquitous self antigen, the cell is eliminated by encountering the antigen early in its development, while survival signals received through the antigen receptor select and maintain a functional lymphocyte repertoire. Adaptive immunity is initiated when an innate immune response fails to eliminate a new infection, and antigen and activated antigen-presenting cells are delivered to the draining lymphoid tissues. When a recirculating lymphocyte encounters its specific foreign antigen in peripheral lymphoid tissues, it is induced to proliferate and its progeny then differentiate into effector cells that can eliminate the infectious agent. A subset of these proliferating lymphocytes differentiate into memory cells, ready to respond rapidly to the same pathogen if it is encountered again. The details of these processes of recognition, development, and differentiation form the main material of the middle three parts of this book.
In Antibody-mediated Immunity Where Does the Antigen First Bind?
Source: https://www.ncbi.nlm.nih.gov/books/NBK27090/
{kind=link}
Post a Comment for "In Antibody-mediated Immunity Where Does the Antigen First Bind?"